传统的药物主要作用于相应的分子靶点 (如激酶、受体、离子通道和转运体等蛋白质靶标)、生物学途径或细胞过程,从而达到治疗疾病的药理作用。小分子化合物和抗体是当前医疗用药的主要形式和药物开发的首选方式。但是只有 10-14% 的蛋白质可以成为小分子化合物或抗体的作用靶点。这限制了小分子化合物和抗体在药物开发中的可能性。
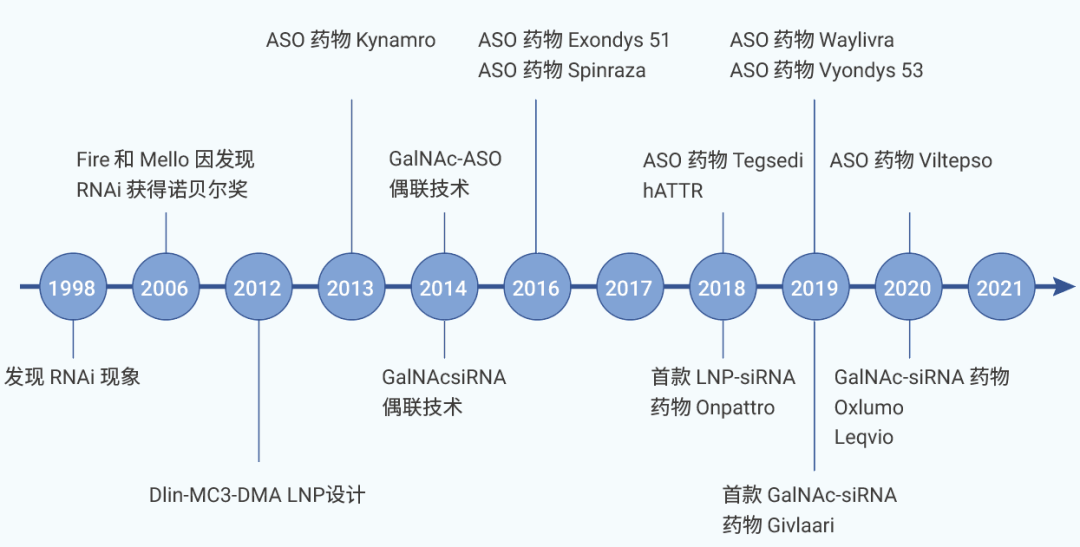
图 1. 小核酸药物的发展历程
传统药物 VS. RNA 药物
然而 RNA 治疗有望扩大药物靶点的范围,包括传统蛋白质和以前无法成为药物靶点的转录本和基因。与其他传统药物相比,核酸药物表现出一些独特的特征 (见图 2)。例如,核酸药物通常以注射的方式给药,而且在体内多次循环使用使得给药频次更低。此外,核酸药物还具有生产制备工艺相对简单和研发周期短 (不需要大规模的药物筛选过程) 的优势。
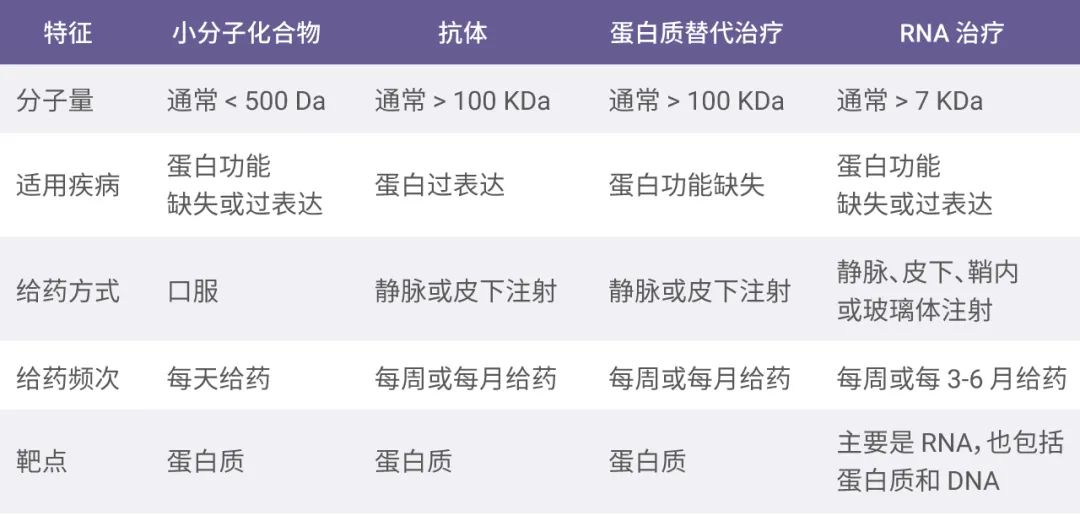
图 2. 小分子化合物、抗体、蛋白质替代治疗和 RNA 治疗的特征
RNA 治疗机制
RNA 治疗包括使用可编码 RNA (如 mRNA) 和非编码 RNA (如小干扰 RNA (siRNA)、microRNA (miRNA)、反义寡核苷酸 (ASO)、核酸适配体 (aptamer)、核酶 (ribozyme) 和引导 RNA (guide RNA (gRNA) 等)。这些核酸具有不同的结构特点和作用机制 。
。
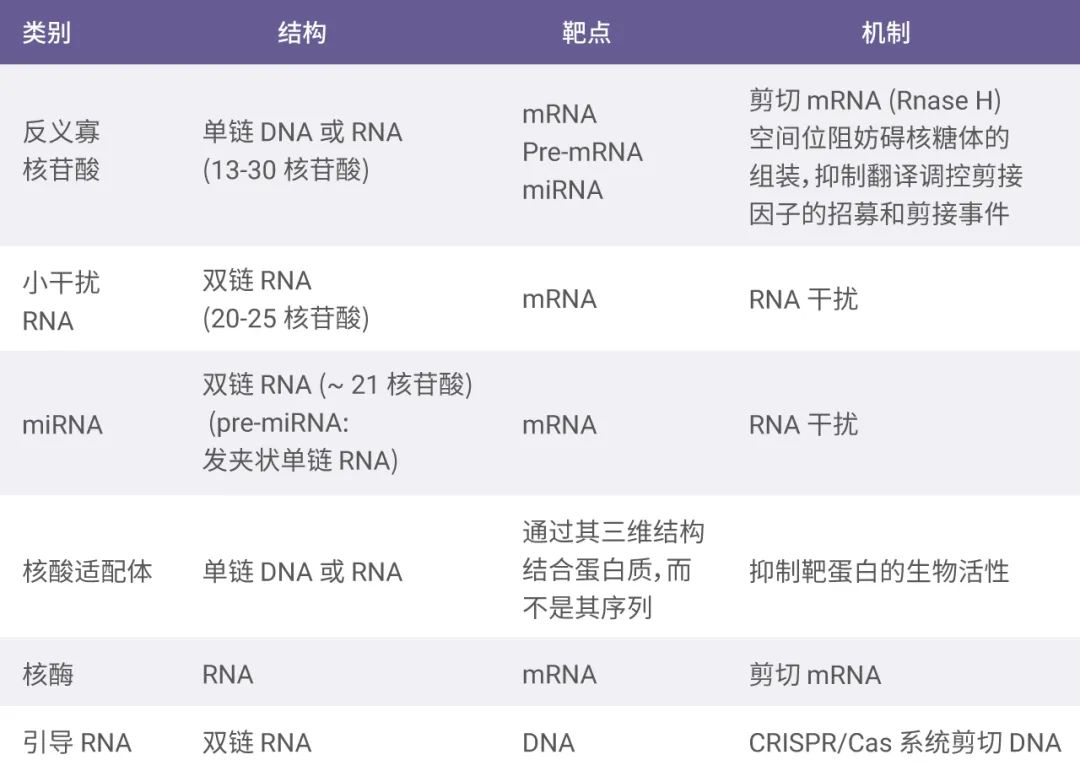
图 3. 不同类型核酸的结构特点和作用机制
ASO、siRNA 和 miRNA 通过碱基互补配对原则靶向 mRNA 或 pre-mRNA,通过基因沉默抑制靶蛋白的表达从而实现治疗疾病的目的。mRNA 分子进入细胞然后翻译成目标蛋白质,可以用于蛋白质替代治疗或疫苗接种。
核酸适配体通过其独特的三级结构而不是其序列与靶蛋白结合。
CRISPR/Cas9 系统由 Cas9 蛋白和引导 RNA 两种分子组成 (“魔剪” CRISPR/Cas9,你 get 到了吗?)。Cas9 蛋白被引导 RNA 分子所激活,发挥识别和切割基因组 DNA 的功能。核酶是具有催化活性的 RNA,可降解特异的 mRNA 序列。
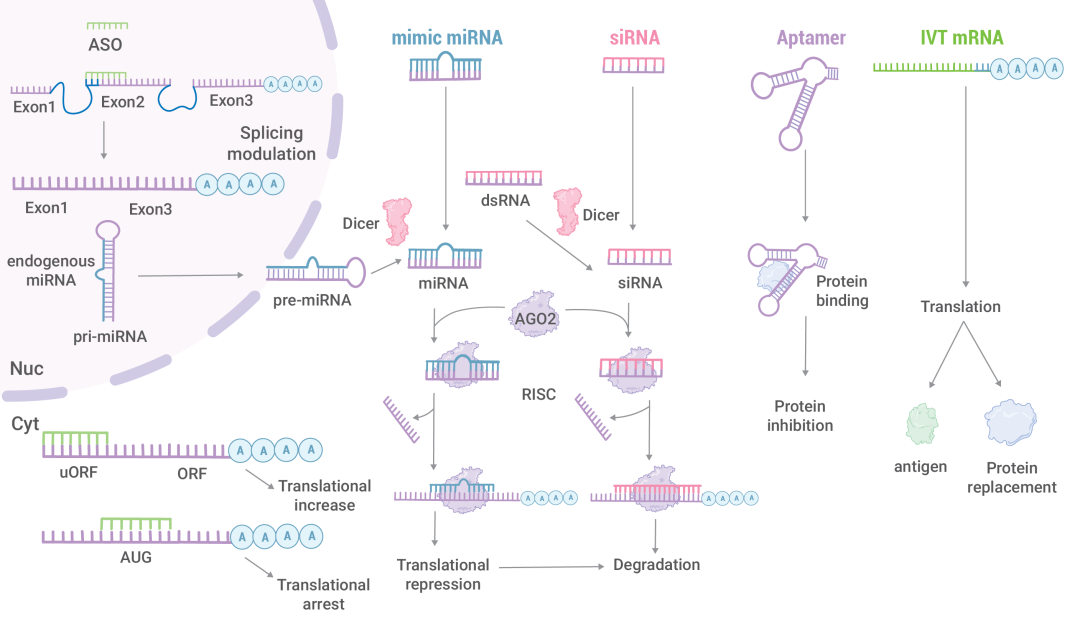
ASOs 可以调节 RNA 拼接,抑制 mRNA 翻译。miRNA 和 siRNA 抑制翻译导致 mRNA 降解。RNA适配体抑制蛋白质活性。IVT mRNA被翻译成宿主蛋白,在细胞内充当抗原或替代蛋白。
核酸药物针对的适应症
利用具有特定序列的核酸作为药物打破了传统药物治疗方法只能作用于靶蛋白的方式。这些核酸药物的候选靶点丰富,适应症分布广。
目前,已获批上市的核酸药物的适应症大多集中于罕见病 (如用于治疗脊髓性肌萎缩症的 Nusinersen (修饰 SMN2 基因的前信使 RNA 剪接,从而促进全长 SMN 蛋白的生成,用于治疗脊髓性肌萎缩症) 。用于治疗急性间歇性卟啉症的 Givosiran,用于治疗 1 型原发性高草酸尿症的 Lumasiran 和用于治疗杜氏肌营养不良症的 Viltolarsen (这几个是已经 FDA 批准的么?
其他用于罕见病治疗的核酸药物在我们之前的文章 “罕见病,其实并不罕见” 中有详细的介绍),后续有望逐渐向肿瘤、神经疾病和代谢疾病等治病领域拓展 (见图 5)。
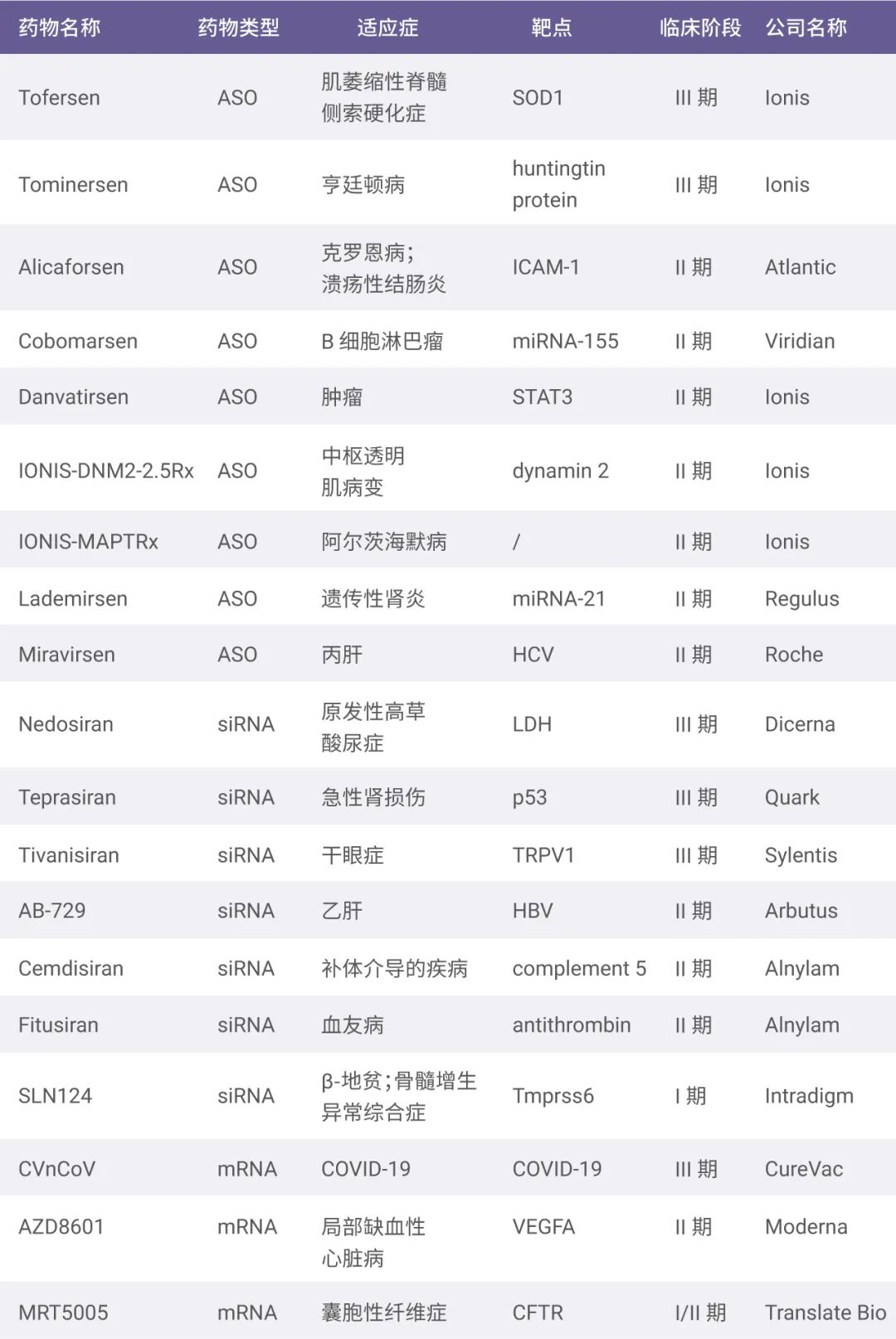
图 5. 处于研发阶段的核酸药物
除了将核酸作为药物用于治疗疾病,科研人员也在尝试开发靶向 RNA 的小分子化合物。RNA 可以被折叠成二级 (如螺旋、茎、环和凸起)、三级 (如结、假结和基序) 和四级 (如复合体) 结构,因此,小分子化合物有可能直接与 RNA 独特的高级结构而不是序列相互作用。以 rRNA 为靶点的新型合成抗生素 (如酮内酯 Telithromycin),靶向核糖体的 Ataluren (用于杜氏肌营养不良治疗),以及靶向 SMN2 pre-mRNA 的 Risdiplam (用于治疗脊髓性肌肉萎缩症) 等药物都已获批用于治疗。
尽管核酸药物具有很好的治疗,但也面临一些挑战。由于核酸通常分子量较大 (单链ASOs ~4 - 10kda,双链siRNA ~ 14kda),且携带负电荷,所以很难有效地进入细胞。此外,裸的核酸在血液中容易被核酸酶降解,还能激活一些免疫识别受体如TLR3/7/8。这些因素限制了核酸药物的的治疗潜力。
为了克服核酸递送的障碍,科学家们已经开发了基于病毒载体和非病毒载体的传递系统(脂质体),以保护寡核苷酸不被降解,最大限度地传递到靶细胞。脂质纳米颗粒通常由阳离子脂质、胆固醇、PEG化脂质和磷脂组成,有助于掩盖核酸携带的电荷,保护其不被核酸酶降解。化学修饰同样可以帮助提高核酸递送的递送效率。2'化学修饰(2'-F、2'-OMe和2'-MOE等)的掺入大大提高了核酸的稳定性和整体半衰期。
不过值得注意的是,与蛋白质不同,RNA 与 DNA 密切相关。
许多最初设计和识别的 RNA 的化合物后来被知道会作用于其他靶点 (例如,直接与 DNA 结合)。因此,在开发 RNA 靶向的小分子化合物时,结构优化是必要的,以提高对 RNA 靶点的选择性。
|
相关产品 |
|
Risdiplam 是一种 SMN2 剪接调节剂,可提高全长 SMN 蛋白的产生水平。 |
|
AB-729 是一种 siRNA,抑制 HBV 病毒复制。 |
|
Cobomarsen 是 miRNA-155 的寡核苷酸抑制剂,抑制与细胞存活相关的多种基因通路 (JAK/STAT,MAPK/ERK 和 PI3K/AKT)。 |
|
Fitusiran 是一种靶向抗凝血酶 mRNA 的 siRNA,降低肝脏中抗凝血酶的产生,可用于血友病的研究。 |
|
IONIS-DNM2-2.5Rx 是一种以 dynamin 2 为靶点的反义药物,可用于中枢透明肌病变 (CNM) 的研究。 |
|
IONIS-MAPTRx 是第一个降低 Tau 蛋白的反义寡核苷酸,可用于阿尔茨海默病的研究。 |
|
Lademirsen 是一种靶向 miRNA-21 的反义寡核苷酸,具有用于 Alport 肾病研究的潜力。 |
|
Miravirsen 是一种靶向 miRNA-122 的反义寡核苷酸,用于 HCV 感染的研究。 |
|
Nedosiran 是一种靶向乳酸脱氢酶 (LDH) 的 siRNA,可用于研究并发终末期肾病 (ESRD) 的原发性高草酸尿症。 |
|
SLN124 是一种靶向跨膜丝氨酸蛋白酶 6 (Tmprss6) 的 siRNA,可恢复铁调素表达和使 β-地贫中铁稳态正常化。 |
|
Teprasiran 是一种小干扰 RNA,可抑制急性肾损伤中 p53 介导的细胞死亡。 |
|
Tivanisiran 是一种靶向 TRPV1 的 siRNA,可用于干眼症的研究。 |
|
Tofersen 是一种靶向超氧化物歧化酶 1 (SOD1) mRNA 的反义寡核苷酸,可用于肌萎缩侧索硬化 (ALS) 的研究。 |
|
Tominersen 是一种靶向亨廷蛋白 (HTT) mRNA 的反义寡核苷酸,可用于亨廷顿病的研究。 |
|
Cemdisiran 是一种 GalNAc 偶联的 siRNA,通过抑制肝脏补体 5 (C5) 蛋白的产生来治疗补体介导的疾病。 |
|
Alicaforsen 是一种 20 个碱基长度的反义寡核苷酸,抑制 ICAM-1 (黏附分子) 的产生。 |
|
Danvantisen 是一种靶向 STAT3 的反义寡核苷酸,具有潜在的抗肿瘤活性。 |
|
是一种可口服的 CFTR-G542X 无义等位基因抑制剂。 |
|
是一种酮内酯,是一种用于治疗呼吸道感染的新型抗生素。 |
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献
1. Damase TR, Sukhovershin R, Boada C, Taraballi F, Pettigrew RI, Cooke JP. The Limitless Future of RNA Therapeutics. Front Bioeng Biotechnol. 2021;9:628137.
2. Mollocana-Lara EC, Ni M, Agathos SN, Gonzales-Zubiate FA. The infinite possibilities of RNA therapeutics. J Ind Microbiol Biotechnol. 2021;48(9-10):kuab063.
3. Feng R, Patil S, Zhao X, Miao Z, Qian A. RNA Therapeutics - Research and Clinical Advancements. Front Mol Biosci. 2021;8:710738.
4. Zhu G, Chen X. Aptamer-based targeted therapy. Adv Drug Deliv Rev. 2018;134:65-78.
5. Damase TR, Sukhovershin R, Boada C, Taraballi F, Pettigrew RI, Cooke JP. The Limitless Future of RNA Therapeutics. Front Bioeng Biotechnol. 2021;9:628137.
6. Yu AM, Choi YH, Tu MJ. RNA Drugs and RNA Targets for Small Molecules: Principles, Progress, and Challenges. Pharmacol Rev. 2020;72(4):862-898.