线粒体质量控制 (mitochondrial quality control,MQC) 是监测线粒体质量的一个综合网络,也是一种细胞内源性保护程序,对于维持线粒体稳态和功能至关重要[1]。MQC 会通过协调生物发生、线粒体裂变、融合、线粒体蛋白水解和线粒体自噬降解等各种过程来共同调控维持线粒体稳态[2]。
通常情况下,线粒体运作一段时间后,功能会呈现下降趋势,这时线粒体间便会发生融合,共用一套内部体系,来维持线粒体正常功能。不久后,线粒体功能再次出现运转障碍,于是线粒体开始分裂,清除损伤线粒体。状态较好的线粒体还将继续融合,被清除的线粒体便会通过自噬途径进行降解回收利用。
线粒体分裂,融合与自噬都是为了让线粒体更好的执行它的任务:线粒体生物发生(图 1)。
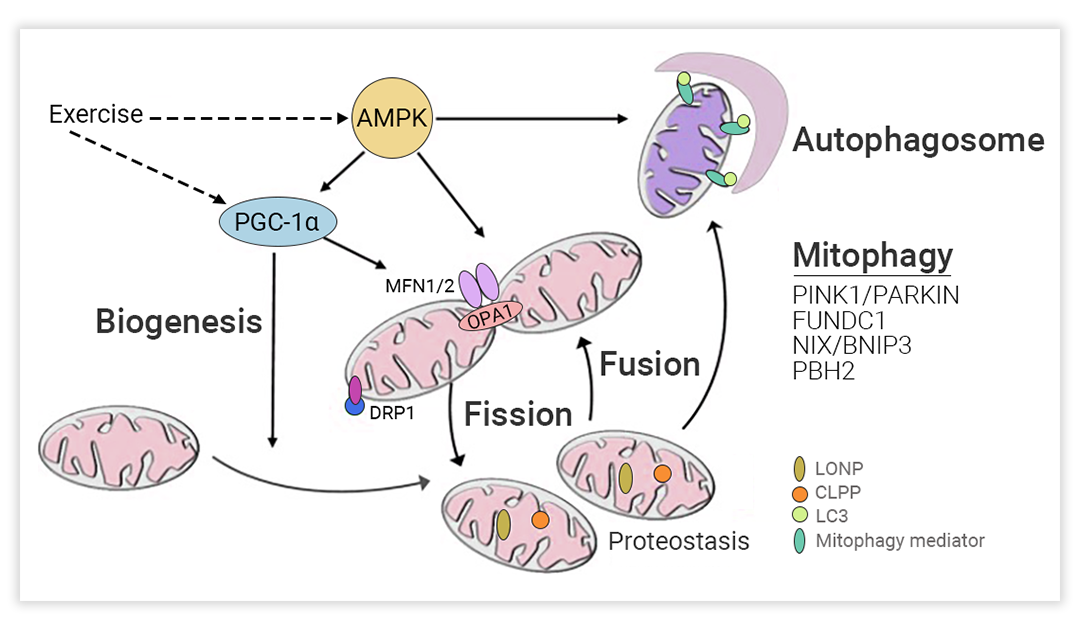
图 1. 线粒体生物发生[1]
线粒体生物发生是一种维持线粒体数量的再生程序,用新的和健康的线粒体替换旧的和受损的线粒体。线粒体生物发生由线粒体基因 (mtDNA) 和核基因 (nDNA) 共同调控,其过程由多个转录因子共同调节发生[3]。通过线粒体生物发生产生的新线粒体可促进 ATP 生成,以满足生理 (如运动和寒冷) 和病理 (如糖尿病缺血性心肌病和缺血性心脏病) 状态下的代谢需求[4]。
线粒体还是一种高度动态的细胞器,可进行连续循环的融合与分裂,以改变线粒体形态、大小及位置,该生理过程被称为线粒体动力学[4]。
融合通常是一种防御反应,两个线粒体可以通过融合它们的内外膜形成一个单一的管状线粒体,来增强线粒体网络的抵抗力 (图 2)[1]。线粒体融合是由线粒体外膜 (OMM) 的线粒体融合蛋白 1 和 2 (Mfn1/2) 和 线粒体内膜 (IMM) 的视神经萎缩蛋白 1 (OPA1) 之间的同型和异型相互作用诱导的。线粒体融合可以决定 mtDNA、膜磷脂和呼吸相关蛋白的交换,以及三羧酸 (TCA) 循环的中间产物。因此,新形成的细胞器具有异质性的膜电位和多样化的蛋白质、代谢物和 mtDNA 池[4]。
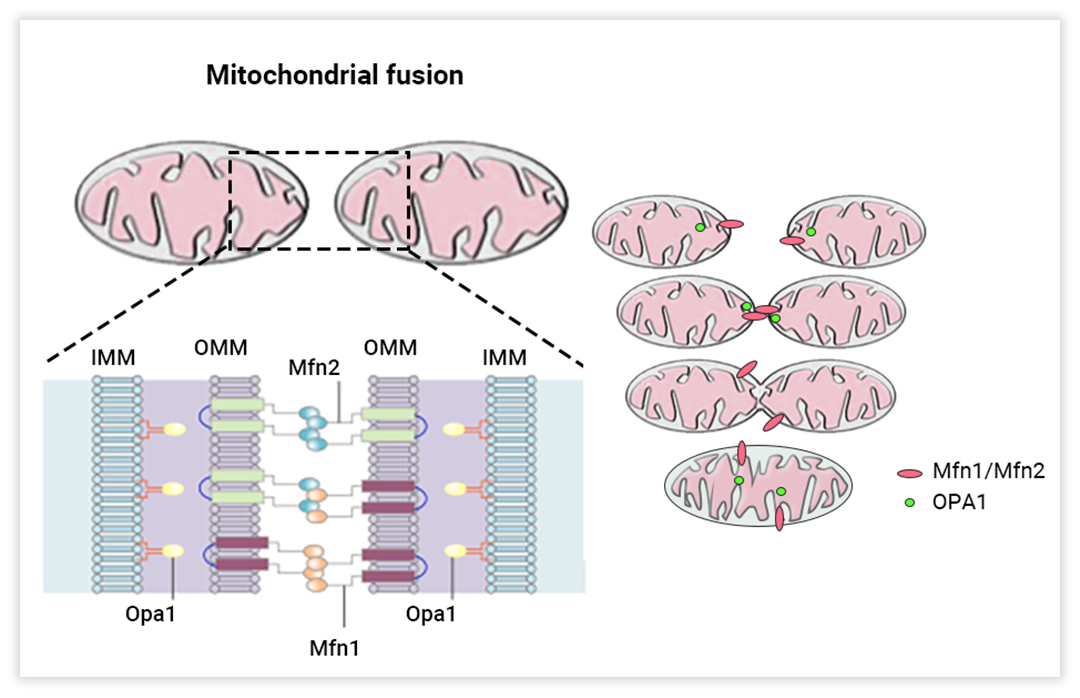
图 2. 线粒体融合的调控机制 (左) 和示意图 (右)[4-5]
相反,线粒体分裂将管状线粒体网络碎片成小的细胞器,有利于通过线粒体自噬消除去极化线粒体。
线粒体分裂主要有三步:1. 动力相关蛋白1 (Drp1) 的磷酸化 (激活);2. Drp1 通过与包括分裂蛋白1 (Fis1)、线粒体分裂因子(Mff) 和线粒体动态蛋白 (MiD49 和 MiD51) 在内的 Drp1 受体相互作用募集到 OMM;3. Drp1 组装成环状结构,环绕并压缩线粒体,消耗 GTP,产生两个单独的细胞器 (图 3)。这一过程不仅可以满足细胞增加的能量需求,还可以从整个线粒体网络中分离出膜电位低的受损线粒体组分,以维持线粒体健康[4]。
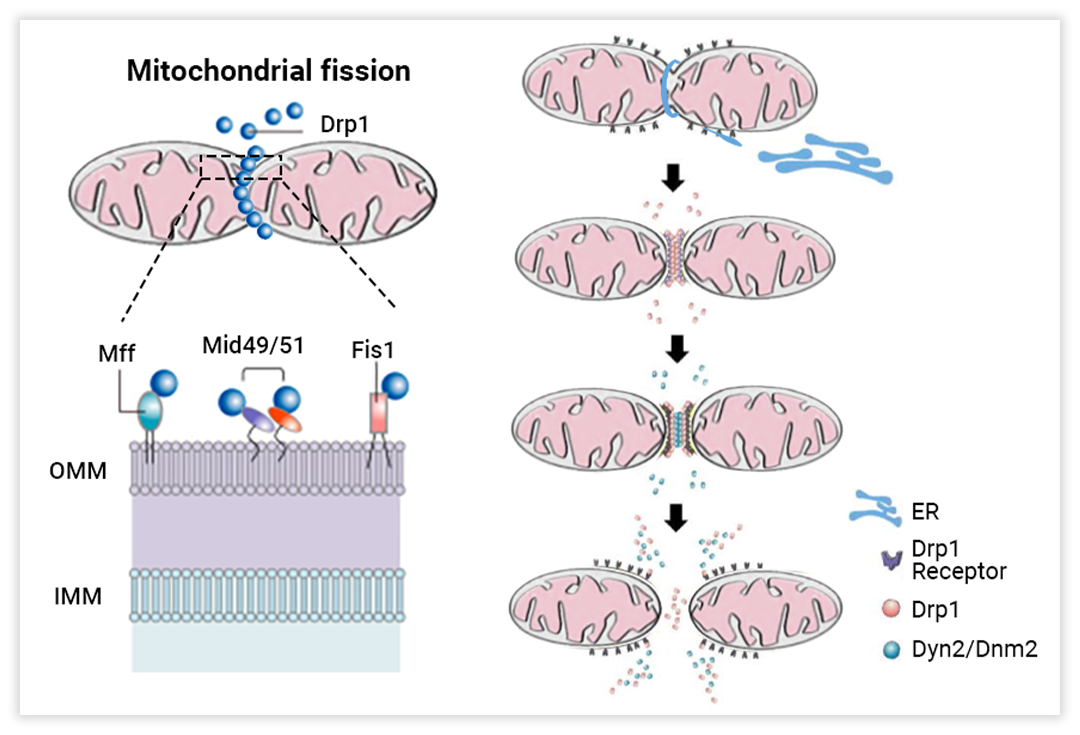
在生理状态下,线粒体融合和分裂互相牵制,使线粒体达到某种动态平衡。该平衡一旦被打破,即线粒体融合和分裂受阻时,将导致线粒体功能受损,最终引发多种疾病[6]。
蛋白水解也是线粒体质量控制机制之一,其对氧化应激、错误折叠、损坏的蛋白质或电子传输链缺陷造成的损害作出反应。提到线粒体蛋白水解,最重要的便是线粒体蛋白酶。
线粒体蛋白酶可降解不同线粒体区室中的缺陷蛋白。在基质中,多种酶共同完成对错误折叠、受损和氧化的蛋白质的降解,以及一些代谢酶的周转,形成 CLPXP 复合物和 Lon 蛋白酶同源物 (LONP)。蛋白质水解产生多肽,这些多肽可以被输出到细胞质或被寡肽酶预序列蛋白酶 (PITRM1) 进一步降解为氨基酸。在膜间隙,HTRA2 降解错误折叠、受损和氧化的蛋白质。
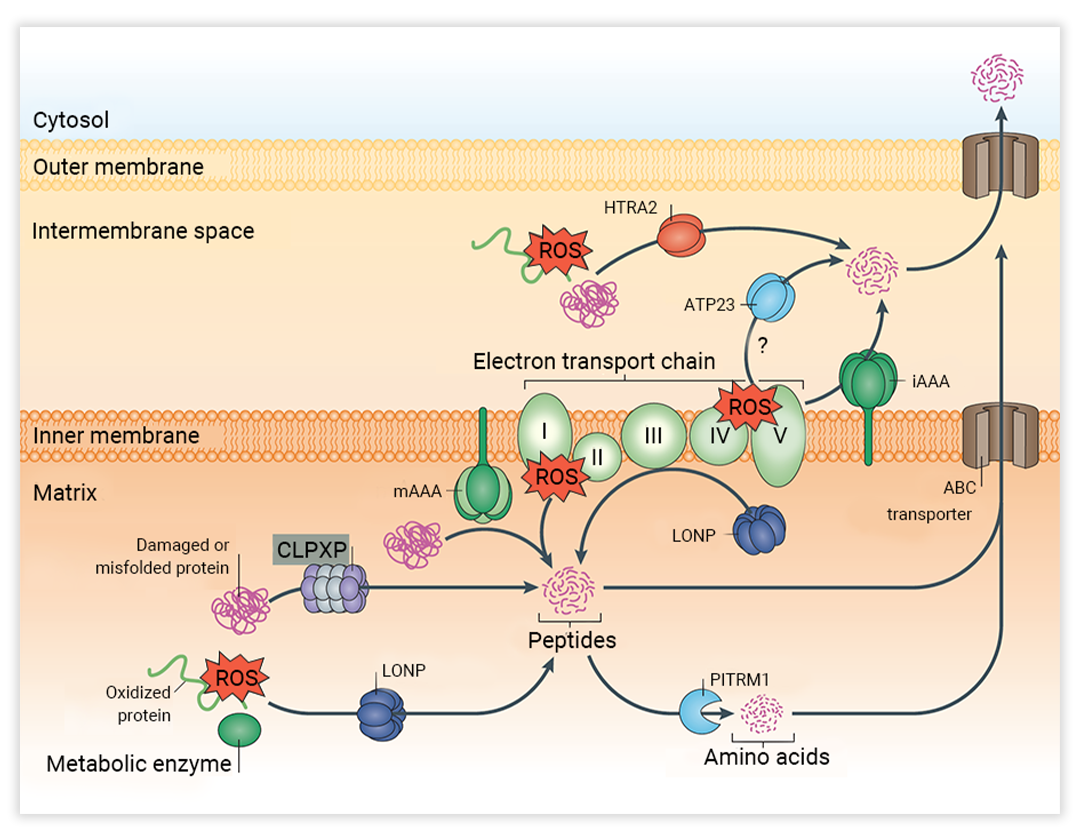
据统计,1,200 种线粒体蛋白质中约有三分之二存在于基质中,主要的基质 AAA 蛋白酶 (如 LONP 和 CLPP) 在线粒体的蛋白质折叠稳态控制机制中起着重要作用,其突变与人类遗传病有关。例如,人类 LONP1 基因的突变会导致 CODAS 综合征[7]。人类 CLPP 基因的突变会导致 Perrault 综合征等。总之,线粒体蛋白水解被认为对维持线粒体功能至关重要。
线粒体自噬降解
往期 小 M 已为大家详细介绍说线粒体自噬,为避免赘述,此处作简要说明。
线粒体自噬主要通过清除受损线粒体来实现质量控制。线粒体自噬最明确的机制之一是 PINK1/Parkin 通路途径,受损的线粒体也可以通过其他自噬途径消除,包括 BCL2 相互作用蛋白 3 样 (BNIP3L) 或 FUN14 结构域包含 1 (FUNDC1) 依赖的线粒体自噬。以及依赖 ULK1 和 ATG5 非依赖的非经典自噬。此外,线粒体也可被 Pink1 - Park2 依赖性线粒体源性囊泡清除。线粒体可能被隔离在空泡中,随后从死亡细胞中挤出。线粒体也可能通过偏头痛体介导的有丝分裂机制分泌[8]。
图 5. 线粒体自噬机制概述[10]
随着研究的深入,越来越多的人类疾病与广泛的线粒体缺陷有关。当线粒体处于过度应激或其质量控制过程失败时,细胞,组织甚至整个有机体都会做出反应,线粒体功能异常将导致代谢、免疫炎症和神经系统疾病的发生[11-12]。
本期小 M 为大家介绍了线粒体相关的质量控制网络,分别从生物发生、线粒体动力学 (裂变/融合)、线粒体蛋白水解和线粒体自噬降解等方面介绍了线粒体的质量控制机制,其共同调控维持线粒体稳态。作为 “科研汪” 的你一定要好好领悟哇~早日发Paper!
| 相关产品 |
| MYLS22 MYLS22 是一种首创的,选择性视神经萎缩 1 (OPA1) 抑制剂。MYLS22 可以靶向内皮细胞 OPA1 减少肿瘤的生长,通过影响 NF-kB 活性和血管生成基因表达抑制血管生成。MYLS22 可抑制线粒体融合,加剧线粒体损伤。 |
| LONP1-IN-2 LONP1-IN-2 (compound 9d) 是一种有效的选择性 LONP1 抑制剂,对 LONP1 和 20 S 蛋白酶体的 IC50 值分别为 0.187 μM 和 >10 μM。LONP1-IN-2 可用于癌症研究。 |
| JC-1 Mitochondrial Membrane Potential Assay Kit 线粒体膜电位检测试剂盒 (JC-1) 是一种以 JC-1 为荧光探针,快速灵敏地检测组织、细胞或纯化的线粒体跨膜电势差的试剂盒,可以用于早期的细胞凋亡检测。 |
| 线粒体靶向化合物库 MCE 收录了 482 种线粒体靶向化合物,主要靶点涉及线粒体代谢、ATP 合酶、线粒体自噬 (Mitophagy)、活性氧 (ROS) 等。MCE 线粒体靶向库是进行线粒体相关药物开发及相关研究的重要工具。 |
| 线粒体保护化合物库 线粒体功能的正常运行不管是在维持细胞正常死亡,还是在线粒体疾病中都有重要意义。MCE 提供 500 个明确或潜在可以保护线粒体的化合物。MCE 线粒体保护化合物库是药物发现和开发的关键。 |
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务
参考文献
[1] Gan Z, et al. Skeletal muscle mitochondrial remodeling in exercise and diseases. Cell Res. 2018;28(10):969-980.
[2] Zhou H, et al. TMBIM6 prevents VDAC1 multimerization and improves mitochondrial quality control to reduce sepsis-related myocardial injury. Metabolism. 2023;140:155383.
[3] Scarpulla RC. Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol Rev. 2008;88(2):611-638.
[4] Zhou H, et al. Role of mitochondrial quality surveillance in myocardial infarction: From bench to bedside. Ageing Res Rev. 2021;66:101250.
[5] Quirós PM, et al. New roles for mitochondrial proteases in health, ageing and disease. Nat Rev Mol Cell Biol. 2015;16(6):345-359.
[6] Archer SL. Mitochondrial dynamics--mitochondrial fission and fusion in human diseases. N Engl J Med. 2013;369(23):2236-2251.
[7] Strauss KA, et al. CODAS syndrome is associated with mutations of LONP1, encoding mitochondrial AAA+ Lon protease. Am J Hum Genet. 2015 Jan 8;96(1):121-35.
[8] Bao F, et al. Mitolysosome exocytosis, a mitophagy-independent mitochondrial quality control in flunarizine-induced parkinsonism-like symptoms. Sci Adv. 2022;8(15):eabk2376.
[9] Nunnari, et al. A mitochondrial protease with two catalytic subunits of nonoverlapping specificities. Science 262, 1997–2004 (1993).
[10] Lu Y, et al. Cellular mitophagy: Mechanism, roles in diseases and small molecule pharmacological regulation. Theranostics. 2023;13(2):736-766.
[11] Farmer T, et al. Tying trafficking to fusion and fission at the mighty mitochondria. Traffic. 2018;19(8):569-577.
[12] Cai Q, et al. Mitophagy in Alzheimer's Disease and Other Age-Related Neurodegenerative Diseases. Cells. 2020;9(1):150. Published 2020 Jan 8.